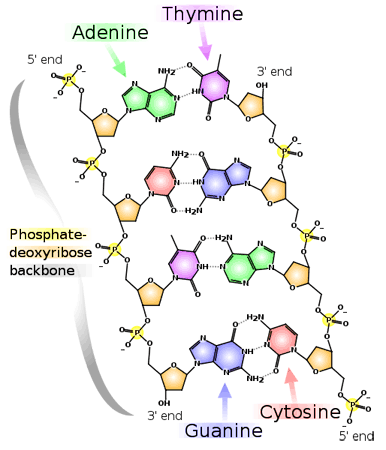
REPLIKACIJA DNK
Replikacija DNK odvija se pre svake ćelijske deobe i omogućava kasniju podelu svakog hromozoma na dve hromatide. Započinje odmotavanjem lanaca DNK i njihovim razdvajanjem. Za oslobođene baze u svakom od lanaca vežu se komplementarne baze koje međusobno poveže DNK polimeraza. Tako na svakom lancu nastane jedan novi lanac i cela se DNK udvostruči.
Sposobnost DNK da duplira samu sebe naziva se replikacija, a od osnovne je važnosti za njenu ulogu naslednog materijala. Za replikaciju je od najveće važnosti princip komplementarnosti vezivanja naspramnih baza u polinukleotidnim lancima DNK. Udvajanju molekula DNK prethodi rasplitanje dvostruke spirale i odvajanje roditeljskih lanaca. Roditeljski lanci služe kao kalup (matrica) za sintezu novog komplementarnog lanca DNK . Svaki novonastali molekul DNK sadrži po jedan stari i jedan novi lanac DNK, pa se zbog kaže da je replikacija semikonzervativan proces(lat. semi= polu; konzervativan=očuvan). Dokaz da je replikacija semikonzervativan proces dobijen je iz eksperimenata sa bakterijom Escherichia coli. Bakterije su gajene na medijumu koji je sadržao teški azot N15 i utvrđeno je da se on ugradio u njihove ćelije i DNK.Zatim su ove bakterije prebačene na medijum koji je sadržao laki azot N14 i u njemu je obavljena samo jedna deoba bakterija. Na osnovu analize sastava DNK utvrđeno je da ona sadrži N15 i N14 u jednakim količinama.
Replikacija se i kod prokariota i kod eukariota vrši bidirekciono, što znači da se od mesta gde počinje vrši istovremeno u oba pravca i to isključivo u 5' - 3' pravcu. Kod prokariota, čija je DNK prstenasta(kružna) replikacija počinje na samo jednom mestu i odvija se bidirekciono. Kružna DNK prisutna je i u eukariotskim organelama: mitohondrijama i hloroplastima, pa se replikacija te DNK vrši slično prokariotskoj.
Kada se lanci DNK razmotaju obrazuje se replikaciona viljuška (ima oblik slova Y). Kod prokariota u svakom trenutku replikacije postoje dve replikacione viljuške :jedna se kreće u smeru kazaljke na satu, a druga u suprotnom smeru. Replikacija DNK u eukariotskim ćelijama započinje istovremeno na mnogo mesta duž hromozoma i teče istovremeno u oba pravca.(Na svakom mestu početka obrazuju se u stvari dve replikacione viljuške.) Na taj način replikacija ukupne hromozomske DNK se u eukariotskim ćelijama završava za nekoliko sati. Brzina replikacije u eukariotskoj ćeliji je 10-100 nukleotidnih parova u sekundi. Ako bi počinjala na jednom mestu,replikacija ukupne lljudske DNK trajala bi oko tri meseca ili više od 500 sati za jedan hromozom.
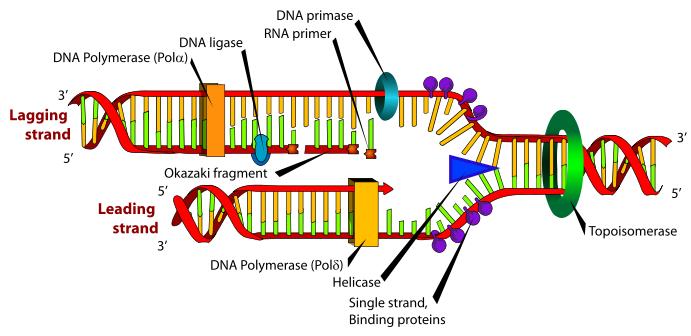
Iako je princip replikacije veoma jednostavan, u ćeliji je to složen proces u kome učestvuje 20-ak enzima i drugih proteina.
Neki od najvažnijih enzima u relikaciji su:
Helikaze su enzimi koji hodaju po DNK raskidajući vodonične veze između lanaca DNK u 5’- 3'’ pravcu. Nukleaze su enzimi koji raskidaju fosfodiestarske veze, pri čemu se razlikuju one koje deluju na krajevima lanaca (egzonukleaze) i one koje deluju na veze unutar lanca (endonukleaze).
DNK- polimeraze su enzimi koji imaju ključnu ulogu u replikaciji. Prokariotske ćelije sadrže tri tipa DNK-polimeraze: I, II i III,pri čemu je DNK-poly III najvažnija jer obavlja najveći deo replikacije; u eukariotskim ćelijama nalazi se najmanje 5 vrsta ovih enzima koje su označene kao DNK polimeraze a, b, g, d i e, pri čemu je najvažnija DNK-poly d. DNK polimeraza III u 5’- 3’ pravcu povezuju nukleotide novog lanca fosfodiestarskim vezama, pošto su se oni postavili komplementarno (A=T, CºG) nukleotidima starog (roditeljskog) lanca. DNK polimeraza (III odnosno d) ima još jednu ulogu: u suprotnom pravcu, 3’- 5’, ona raskida fosfodiestarske veze između pogrešno vezanih nukleotida novog lanca. Ukoliko DNK polimeraza naiđe na pogrešno sparen nukleotid, ona upotrebi svoju egzonukleaznu aktivnost u smeru 3’- 5’ i raskine vezu tog nukleotida sa lancem. Ta njena egzonukleazna aktivnost omogućava ispravljanje grešaka, koje tokom replikacije nastanu što doprinosi tome da je replikacija izuzetno tačan proces.Greške pri ugrađivanju nukleotida javljaju se sa učestalošću od 10-10.
Ligaze deluju suprotno nukleazama – one delove novog lanca povezuju u celinu obrazujući između tih delova fosfodiestarske veze. Primaza međusobno povezuje nukleotide RNK u kratke lance da bi time omogućila dejstvo DNK polimeraze. (dejstvo ovog enzima biće naknadno još objašnjeno).
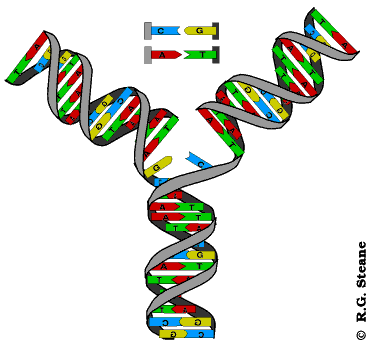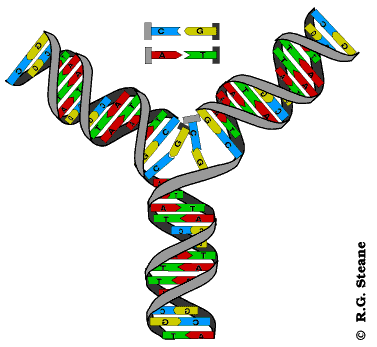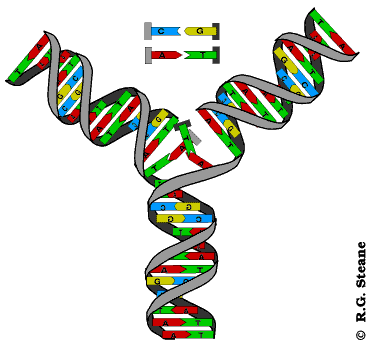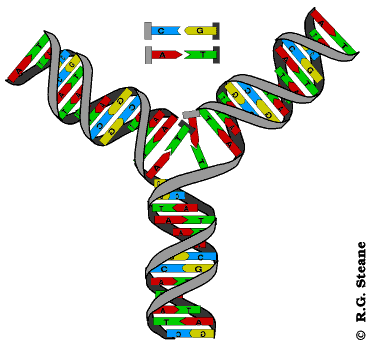
Dvolančana zavojnica DNK je vrlo stabilna struktura, tako da su ćeliji neophodni mehanizmi koji će omogućiti razdvajanje lanaca i formiranje replikativne viljuške. U ovim mehanizmima učestvuju helikaze i još neki proteini. Endonukleaza zasijeca samo jedan lanac DNK čime počinje njeno rasplitanje. Helikaze raskidaju vodonične veze između lanaca čime se lanci razdvajaju i obrazuju se replikativne viljuške. Pošto se replikacija odvija bidirekciono, na svakom mestu gde ona počinje obrazuju se dve replikativne viljuške koje se kreću u suprotnim smerovima. Pratićemo sada šta se dešava u jednoj replikativnoj viljušci. Svaka r. viljuška je asimetrična jer su lanci antiparalelni, a istovremeno se naspram oba sintetišu novi lanci u 5’- 3’ pravcu. Lanac DNK koji se sintetiše u pravcu kretanja replikativne viljuške naziva se vodeći lanac, a onaj koji se sintetiše u suprotnom smeru je lanac koji zaostaje. Vodeći lanac se sintetiše kao celovit dok se ovaj drugi sintetiše u vidu delova koji se nazivaju Okazakijevi fragmenti. Okazakijevi fragmenti su naziv dobili prema autoru koji ih je otkrio; imaju dužinu od 1000-2000 nukleaotida u prokariotskim, odnosno 100-200 nukleotida u eukariotskim ćelijama; naknadno se spajaju ligazama pa pošto se sinteza tog lanca završava sa zakašnjenjm on dobija naziv lanac koji zaostaje.
Sintezu oba lanca obavlja DNK polimeraza tek pošto se veže za roditeljski lanac koji služi kao matrica. Ovaj enzim ne može da se veže za ogoljeni lanac-matricu već zahteva postojanje začetnika(prajmera; engl. primer). Začetnik je kratki lanac RNK i njegovu sintezu katalizuje enzim primaza. Kada se kratki lanac RNK komplementarno spari (hibridizuje) sa početkom lanca matrice to omogućuje vezivanje DNK polimeraze i počinje sinteza novog lanca. Za sintezu lanca koji zaostaje potrebno je da se sintetiše veći broj začetnika. Okazakijeve fragmente, po završetku sinteze, međusobno povezuje enzim ligaza.
 |
 |
 |
 |
 |
 |