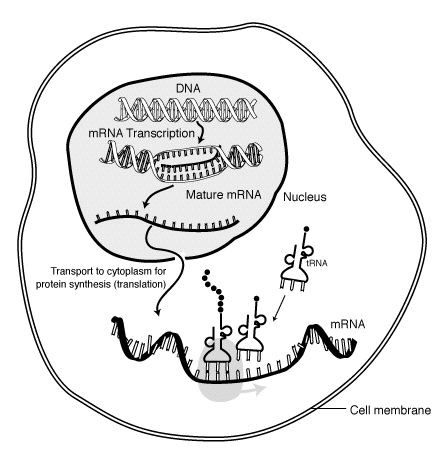
BIOSINTEZA PROTEINA
Genetička informacija se odvija kroz dvije složene etape : Transkripcija (lat. transcripcio - prepisivanje) i Translacija (lat. translacio - prevođenje).Transkripcija je sinteza RNK kao kopije dela jednog lanca DNK (gena) koju katalizuje enzim RNK polimeraza. Vrši se u jedru eukariota u 5'-3' pravcu. Da bi se izvršila transkripcija (prepisivanje) lanci DNK moraju da se razdvoje i jedan od njih služi kao kalup prema kome se ređaju komplementarni nukleotidi RNK : naspram adenina postavlja se uracil, a naspram guanina – citozin i obrnuto. Transkripcija DNK je ključni korak u prenošenju informacije od DNK do proteina. Sinteza i-RNK počinje kada je ćeliji potreban određeni protein.
Za aktivnost RNK-polimeraze, slično kao i za DNK-plimerazu, potrebno je prisustvo DNK lanca-matrice. Međutim, za razliku od DNK-polimeraze, za RNK polimeraze nisu potrebni prajmeri. U principu, bilo kojii od dva lanca DNK može da posluži kao matrica za sintezu RNK, ali se u toku transkripcije prepisuje samo jedan. Naspram matičnog lanca DNK po principu komplementarnosti mogu se sintetisati tri osnovne vrste RNK: iRNK, tRNK i rRNK. Novosintetisani lanac RNK je uvek i po smeru i po redosledu nukleotida identičan onom lancu koji nije prepisivan (samo što umesto T sadrži U) i naziva se smisleni (engl. sense) lanac.
![]()
Transkripcija se odvija kroz tri faze:
U fazi inicijacije RNK polimeraza se vezuje za mesto na genu nazvano promotor. Promotor sadrži mesto za vezivanje RNK polimeraze i on se ne prepisuje u procesu transkripcije. Promotor sadrži kratki niz nauzmenično vezanih T i A uzvodno od mesta početka transkripcije; ovaj se niz zbog karakterističnog rasporeda parova A=T naziva TATA blok; 85% eukariotskih gena u svojim promotorima sadrži TATA blok. U fazi elongacije RNK polimeraza odvija jedan zavoj DNK pri čemu se lanci razdvajaju. Jedan od lanaca služi kao matrica prema kome RNK polimeraza ređa komplementarne nukleotide RNK u 5'-3' pravcu.. Ređanje nukleotida RNK vrši se sve dok RNK polimeraza ne stigne do niza nukleotida koji se nazivaju terminacioni (stop) signal. Najčešći stop-signal je niz od nekoliko uzastupno poređanih adenina (AAAA...). Na tom mestu se transkripcija zaustavlja, a novonastali molekul RNK se oslobađa sa matrice ( do tada je bio za matricu vezan samo svojim 3' krajem) . To predstavlja poslednju fazu terminacije. Kod prokariota, čiji geni nemaju introne, prepisana RNK (primarni transkript) predstavlja zrelu RNK koja se odmah može uključiti u sintezu proteina. Kod eukariota se, međutim, transkripti moraju u jedru dodatno obraditi da bi postale zrele RNK sposobne da otpočnu sintezu proteina. Tek tako obrađene šalju se u citoplazmu gde će se obaviti sinteza proteina.
Dodatna obrada prepisanih RNK sastoji se u:
Mehanizam iskrajanja introna mora biti takav da obezbedi maksimalnu preciznost, jer bi greška u samo jednom nukleotidu potpuno promenila sve kodone u i-RNK od tog mesta nadalje i proizvela pogrešnu informaciju za sintezu proteina. Sinteza proteina se obavlja u procesu nazvanom translacija u kome se abeceda nukleinskih kiselina sa četiri slova (A; G;C;T) mora prevesti na jezik proteina sa 20 različitih aminokiselina. Dok se replikacija i transkripcija eukariota vrše u jedru, translacija se odvija na ribozomima u citoplazmi. Translacija je proces kojim se niz nukleotida u i-RNK prevodi u niz aminokiselina u proteinu. Ulogu prevodioca u ovom procesu (translatio=prevođenje ) igraju t-RNK koje jednom svojim krajem (antikodonom) iščitavaju kodon na i-RNK, a na drugom kraju nose odgovarajuću aminokiselinu.
Da bi se mogli razumeti procesi koji se vrše za vreme translacije, potrebno je upoznati strukturu i osobine t-RNK. T-RNK su najmanje veličine među RNK molekulima (veličina im je svega 70-90 nukleotida). Njihova sekundarna struktura ima oblik deteline sa tri lista i obrazuje se vodoničnim vezama između komplementarnih baza (samo što za razliku od DNK ovde te baze pripadaju istom lancu). Molekul t-RNK sadrži 4 kraka, od kojih tri imaju po jedan dvolančani deo (dršku) i jedan jednolančani deo (petlju). Krak koji ne sadrži petlju predstavlja mesto vezivanja aminokiselina. Na njegovom 3' kraju nalazi se triplet CCA za koji se vezuje aminokiselina. Naspram ovog kraka nalazi se krak koji nosi antikodon. Vezivanje aminokiseline za t-RNK katalizuje enzim aminoacil-t-RNK sintetaza koji omogućava da se ova veza obogati energijom oslobođenom razlaganjem ATP-a. Ta energija će kasnije biti upotrebljena za formiranje peptidne veze. Prema tome, t-RNK imaju veoma značajnu ulogu u prevođenju genetičke informacije sa jezika nukleotida na jezik aminokiselina. Ispravno čitanje genetičke informacije (tačnost translacije) u velikoj meri zavisi od toga da se svaka aminokiselina veže baš za odgovarajuću t-RNK, kao i od pravilnog vezivanja kodona i antikodona (A=U; GºC).
![]()
Translacija se odvija kroz tri faze: inicijaciju, elongaciju i terminaciju.
Prva faza, inicijacija, obuhvata procese kojima se vezuje i-RNK sa malom i velikom subjedinicom ribozoma. U ribozomu ima dva mesta, tzv. P i A mesto (P-peptidil mesto; A-aminoacil mesto, P mesto je ono na kome je vezana t-RNK sa povezanim aminokiselinama, a A mesto je ono na koje dolazi t-RNK sa novom aminokiselinom), za koje se mogu vezati dve t-RNK. Prva t-RNK koja ulazi u ribozom je ona sa antikodonom koji je komplementaran start kodonu AUG (kod prokariota pored AUG, start kodon može da bude i GUG). Ta t-RNK nosi aminokiselinu metionin i vezuje se za P mesto. Na A mesto u ribozomu dolazi druga t-RNK čiji antikodon je komplementaran narednom kodonu i-RNK (prvi posle start kodona).Dve aminokiseline su sada blizu jedna drugoj. Kida se veza između metionina i t-RNK, a oslobođena energija se koristi za stvaranje peptide veze između dve prve aminokiseline. Prva t-RNK napušta ribozom, a druga po redu t-RNK prelazi sa A na P mesto čime se ribozom pomera za jedan kodon duž i-RNK. Ribozom se pomera u 5'-3' pravcu. Sada se na P mestu nalazi t-RNK za koju su vezane dve aminokiseline, a A mesto je slobodno. Na A mesto ulazi sledeća po redu t-RNK koja nosi antikodon komplementaran trećem kodonu i-RNK i proces se ponavlja sve dok se u i-RNK ne stigne do stop kodona. Pošto stop kodon ne određuje ni jednu aminokiselinu, za njega se vezuje neki od proteinskih faktora (RF1, RF2 ili RF3 kod prokariota, dok je kod eukariota samo jedan faktor označen kao eRF; RF-faktor relaksacije) što dovodi do raspada čitavog kompleksa (odvajaju se subjedinice ribozoma od t-RNK i i-RNK i oslobađa se novosintetisani polipeptidni lanac).
![]()
 |
 |
 |
 |
 |
 |